
Introduction
|
Partie IV: Niveau génétique Allons plus loin pour découvrir les mécanismes génétiques de l’horloge biologique. Expérience: En 1970, ŕ l’Institut de technologie de Californie, Ron Konopka et Seymour Benzer, ont réalisé leurs études sur des drosophiles. Ils ont observé 2000 mouches élevées dans 2 incubateurs ŕ l’intérieur desquels la lumičre suivait un rythme périodique (lever et coucher de soleil), et oů l’on introduisait de la nourriture et de l’air. Résultat: la plupart des animaux gardaient un rythme circadien normal de 24 heures avec 12 heures d’activité et autant de repos ; mais 3 mouches développčrent un comportement anormal dű ŕ des mutations, qui se traduisit soit par un cycle de 19 heures, soit un de 28 heures, et męme par l’absence de cycle. Quels étaient les gčnes endommagés? En 1984, 3 équipes identifient le gčne muté : il s’agit du gčne per (period) situé sur le chromosome X. Les 3 mouches possédaient 3 altérations différentes de ce męme gčne. Ces différentes mutations provoquent selon les cas un raccourcissement ou un rallongement de la période des rythmes physiologiques ou męme leur abolition. On s’est aperçu qu’au cours d’un cycle 12 heures de lumičre/12 heures d’obscurité, l’ARNm de per est présent en faible quantité au début du jour et ŕ un taux trčs élevé au début de la nuit. Ce gčne détermine la périodicité circadienne et la valeur exacte de cette période. Agit-il seul? L’étude se porta sur l’observation de 7000 mouches. Une mouche dépourvue de rythme circadien fut isolée. La mutation était située sur le chromosome 2 : découverte du gčne tim (timeless) en 1994. Son expression suit les męmes variations que per. A partir des gčnes per et tim, on synthétisa les protéines PER et TIM. On remarqua ensuite que, réunies dans une męme solution, ces 2 protéines se liaient. Chez les drosophiles mutantes tim (qui ne peuvent donc plus fabriquer la protéine TIM), les oscillations circadiennes de l’ARNm de per sont supprimés ; inversement si l’on empęche l’organisme de produire la protéine PER. En effet PER et TIM, lorsqu’elles atteignent un niveau élevé dans le cytoplasme, s’associent en dimčre, transféré dans le noyau qui inhibe l’expression des gčnes per et tim. Au bout de quelques heures, les enzymes dégradent les dimčres PER-TIM et les gčnes peuvent ŕ nouveau produire leur protéine : un nouveau cycle commence. En 1997, les équipes de Hajime Tei (Université de Tokyo), de Hitoshi Okamura (Université de Kobe), et de Cheng Chi Lee (Collčge de médecine Baylor), isolčrent les analogues du gčne per chez la souris et l’homme. Un an plus tard, on trouva les analogues du gčne tim. Une importante découverte suivit : ces gčnes présents dans toutes les cellules s’expriment dans le noyau suprachiasmatique. Comment sont-ils activés? En 1997, Joseph Takahashi et ses confrčres de l’Institutmédical Howard Hugues, isolent chez la souris, la drosophile et encore bien d‘autres mammifčres, le gčne clock, qui, lorsqu’il est muté supprime le rythme circadien. D’aprčs sa structure, la protéine CLOCK est un facteur de transcription. Les expériences réalisées chez les drosophiles et les souris ont confirmé l’hypothčse selon laquelle CLOCK, associée ŕ une protéine appelée CYCLE chez la drosophile et BMALI chez la souris, stimule la transcription des gčnes per et tim. Depuis peu, Steeve Reppert et l’Université de Haward, explorent des signaux spécifiques qui relient les horloges biologiques de la souris et de divers comportement modulés par ces horloges. L’hypothčse actuellement retenu par les scientifiques est donc que l’inhibition de la transcription de per et tim par le dimčre PER-TIM s’effectue via l’inhibition de CLOCK. De plus, selon une étude menée en 1998, le dimčre PER-TIM peut aussi stimuler la transcription du gčne clock. Mais pour l’heure, les scientifiques ne savent pas comment PER-TIM peut jouer deux rôles assez différents. Les 4 gčnes vus précédemment constituent le coeur de l’horloge biologique des mouches et avec quelques modifications celle de la plupart des animaux. Mais il semble difficile d’imaginer que seuls 4 protéines PER, TIM, CLOCK et CYCLE gčrent tous les mécanismes de l’horloge biologique. En 1998, aprčs la découverte de la protéine DOUBLETIME, le systčme s’est révélé plus complexe. DOUBLETIME est une kinase, une protéine qui, par une réaction de phosphorylation, peut ajouter un groupement phosphate sur d’autres protéines. Ainsi DOUBLETIME phosphoryle PER ce qui la rend instable et l’empęche de former un dimčre avec TIM. On a aussi découvert la protéine CRY. Cette protéine appartient ŕ la famille de cryptochromes, molécules réceptrices de lumičre. Chez la drosophile, CRY, exposée ŕ la lumičre, se lie ŕ TIM alors rendue incapable de former un dimčre avec PER. En somme, DOUBLETIME et CRY en agissant modifient la disponibilité des 2 protéines centrales de l’horloge. De plus, CRY n’a pas la męme fonction chez la souris que chez la drosophile. En effet, chez la souris, CRY se substitue ŕ TIM pour former un dimčre PER-CRY fonctionnellement identique au dimčre PER-TIM de la drosophile. En fait, CRY se trouve selon que l’on étudie la drosophile ou la souris dans 2 rôles opposés : activateur et inhibiteur de la transcription de per. Encore plus petit? Le gčne per est subdivisé en per1, per2, et per3. L’expression de ces 3 gčnes est commandée par le noyau suprachiasmatique. D’aprčs Norio Ishida ŕ l’Institut Tsukuba, on sait que le gčne per2 est exprimé dans les tissus périphériques tel le foie, les poumons, les reins avec un rythme différent selon les tissus. Selon Gijsbertus Van der Horts (Université de Rotterdam), 2 protéines nommées cry1 et cry2 seraient synthétisées dans certaines cellules de la rétine et jouerait un rôle important dans la régulation de l’horloge biologique. Et ailleurs...? Plusieurs autres études ont montré que le cerveau n’était pas le seul responsable des rythmes circadiens. Les scientifiques se sont aperçus que lorsqu’ils mettaient en culture un organe de la drosophile, les cellules de celui-ci gardaient un rythme circadien. On a donc admis la présence d’horloges périphériques chez la drosophile (antennes, région buccale, pattes, ailes, tubules malpighiens...). Mais peut-on affirmer la męme chose chez les vertébrés ? On a prouvé que oui. En effet, on observe des rythmes circadiens dans les organes isolés comme par exemple chez le poisson zčbre, petit poisson couramment utilisé par les généticiens. Chez lui l’expression du gčne homologue ŕ clock oscille de façon circadienne dans presque tous les tissus. On sait également que le coeur et le rein mis en culture (séparés du reste du corps) manifestent des rythmes circadiens pendant plusieurs jours. Ce résultat qui peut ętre étendu ŕ toutes les cellules du poisson zčbre prouve donc que les tissus peuvent agir indépendamment. De plus, les rythmes de ces organes ex vivo ou de ces cellules sont régulés par la lumičre ce qui amčne ŕ penser qu’il existe des photorécepteurs dans ces tissus. Les scientifiques ne les ont malheureusement pas encore découverts. Quant aux mammifčres il semble, d’aprčs des expériences faites sur des rats, qu’ils ont eux aussi des horloges périphériques. A l’heure actuelle les scientifiques cherchent encore le rôle physiologique de ces horloges périphériques. Fonctionnent-elles vraiment indépendamment des horloges centrales ou ŕ l’inverse sont-elles de simples oscillations coordonnées par ses derničres. Le débat reste ouvert. Notons que récemment on s’est aperçu que les drosophiles auxquelles on avait supprimé les horloges périphériques ne manifestaient plus d’oscillation de leur capacité olfactive. Existe-t-il chez l’homme un lien direct entre gčnes de l’horloge biologique et pathologie ? D’aprčs une étude récente, un nucléotide du gčne clock humain varie d’un individu ŕ l’autre et cette variation semble liée avec le temps du sommeil. Des études sont encore en cours. 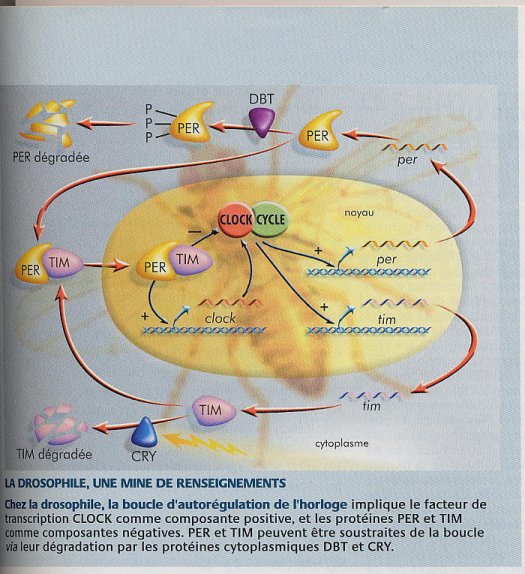
|